Vi presentiamo il post di @vinamra intitolato Bacterial Microcompartments: Nano-Bioreactor premiato dal DaVinci.Times team come miglior post della settimana.
The translation of the post was made by @aditili and has been pre-checked by the @davinci.times team.
Microcompartimenti batterici: Nano-Bioreattore
Salve Steemians!
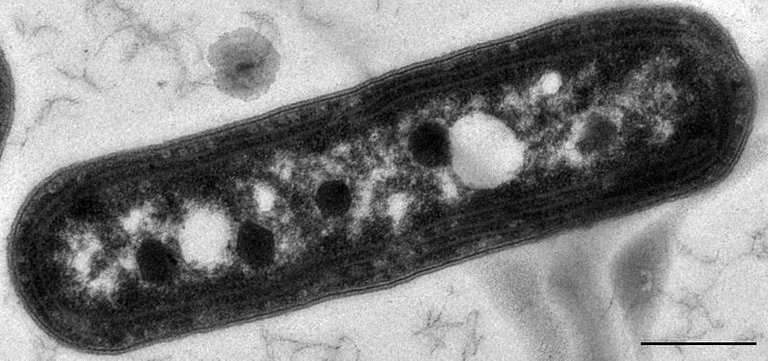
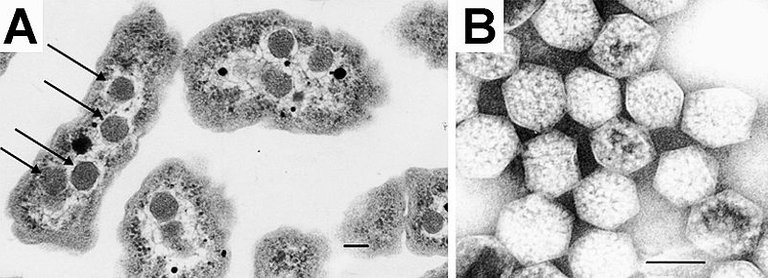
Nell'articolo di oggi parlerò dei microcompartimenti batterici (BMC): essi sono simili agli organuli della cellula eucariota, struttura che racchiude proteine ed enzimi all'interno di una parete proteica. Il primo microcompartimento fu scoperto nel 1950 usando la microscopia elettronica nel citoplasma dei cianobatteri. Si è scoperto che le strutture erano molto piccole nel range di 100-200 nm di diametro, prima si pensava che si trattasse di virus ma in seguito studi con Thiobacillus è stato segnalato come guscio proteico riempito con RuBisCO (Ribulosio Bisfosfato Carbossilasi/Ossigenasi ) e svolge la funzione di fissazione del biossido di carbonio attraverso il ciclo di Calvin (1). La parete proteica è di natura molto selettiva e consente solo l'ingresso di alcuni substrati. La separazione degli enzimi dall'ambiente del citosol impedisce agli enzimi di metabolizzare il metabolismo tossico e riduce il rischio di interruzione da reazioni indesiderate (2).
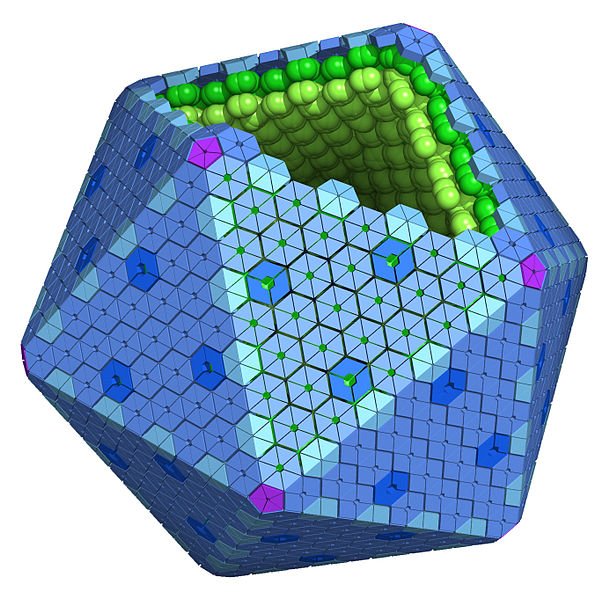
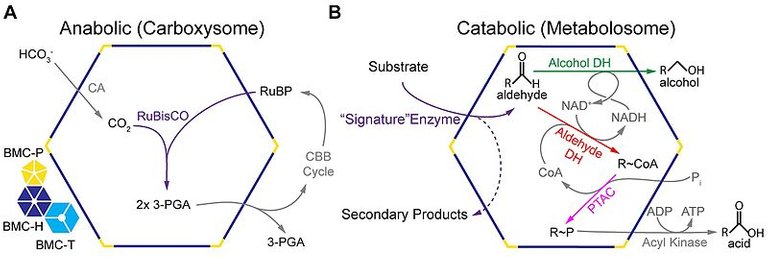
I microcompartimenti batterici sono di due tipi, anabolizzanti e catabolici, in base alla funzione che svolgono. L'unico BMC noto per eseguire il processo anabolico è il carbossisoma presente nei cianobatteri. I Carbossisomi BMC contengono enzimi RuBisCO (che fissano l'anidride carbonica per creare due molecole di fosfoglicerato) e anidrasi carbonica (che catalizza la conversione di bicarbonato in biossido di carbonio) per cui avviene la fissazione del biossido di carbonio attraverso il ciclo di Calvin (3). I BMC catabolici o metabolosomi, si esprimono solo in presenza del substrato. Le BMC sono quindi denominate sulla base del substrato metabolizzato, ad esempio nell'utilizzo di propanediolo (PDU-BMC), di etanolamina (EUT-BMC) di fucosio e di ramnosio (4). In essi il nucleo enzimatico rimane comunque simile e si caratterizza per un enzima di firma (specifico per il substrato iniziale), aldeide deidrogenasi, alcol deidrogenasi e fosfotransacilasi. (5).
Come precedentemente accennato, i BMC risultano selettivamente permeabili a determinati composti. Ad esempio, nel caso dei carbossisomi, la parete BMC è permeabile a RuBP, bicarbonato e fosfoglicerolo, mentre l'ingresso di ossigeno e anidride carbonica è limitato (6). Un altro esempio è la permeabilità al propanediolo, al propanolo ed alla vitamina B12, mentre non lo è alla propionaldeide per prevenire il danno cellulare (7).
Si suppone che la natura selettiva delle proteine del guscio BMC sia dovuta alle cariche delle proteine sulle pareti guscio. I pori dei carbossisomi infatti hanno cariche positive globali che si traducono in ingresso di ioni bicarbonato caricati negativamente (8).
Parliamo della struttura del guscio BMC e del loro assemblaggio in carbossisomi e metabolosomi. I principali elementi strutturali delle proteine glicemiche sono: BMC-H, BMC-T e BMC-P.
Il BMC-H è il principale elemento proteico per formare un homoesamero ciclico con un lato convesso e un lato concavo e contiene un singolo dominio Pfam00936 (9).
Il successivo è il BMC-T, simile allo pseudoesamero ma è un trimero determinato dalla fusione di due domini Pfam00936.
Attraverso l'allineamento di BMC-H e BMC-T si forma un poro sull'asse centrale di simmetria che funge da canale per i metaboliti e permette di poter entrare e uscire dal guscio
(10). BBMC-H e BMC-T danno forma a nuovi livelli di guscio per cui è necessaria una proteina pentamerica per tappare i vertici del guscio.
Ecco che dunque il BMC-P forma tale struttura pentamerica assemblando un singolo dominio Pfam03319 (11).
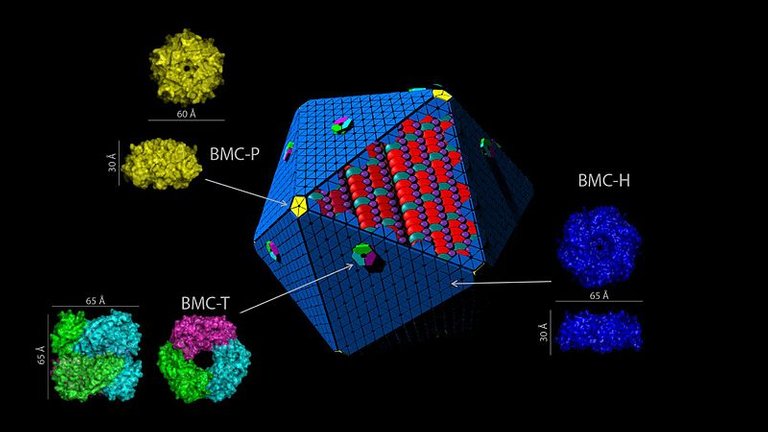
Così gli scienziati hanno iniziato a interessarsi allo studio dell'assemblaggio di BMC nei carbossisomi e nei metabolosomi. I carbossisidi sono divisi in due tipi sulla base del loro modo di assemblarsi: α-carbossisomi e β-carbossisomi. Nel primo tipo nucleo e guscio vengono assemblati di pari passo, nel secondo invece viene assemblato per primo il nucleo e successivamente il guscio (12). L'analisi filogenetica della proteina BMC-P ha mostrato la sua somiglianza con i β-carbossisomi sebbene le vie di assemblaggio dei metabolosomi non siano ancora state spiegate sperimentalmente (2).
Parliamo adesso delle applicazioni biotecnologiche dei microcompartimenti batterici. Essi possono infatti essere utilizzati nei nanobioreattori. Obiettivo principale dell'ingegneria metabolica è di progettare vie metaboliche che abbiano un'alta resa ai prodotti chimici e farmaceutici. Molti fattori come accumulo di sostanze chimiche intermedie e prodotti inibitori condizionano in negativo la resa. Tuttavia in natura queste condizioni possono essere aggirate grazie alla compartimentazione, come avviene ad esempio nei perossisomi (13). Gli scienziati hanno iniziato a utilizzare impalcature di proteine o di DNA per circoscrivere gli enzimi entro un compartimento ed aumentare la resa del prodotto, come ad esempio avviene nella produzione di acido glucarico e mevalonato (14). Un altro utilizzo che può avvenire è relativo al deposito di nanoparticelle metalliche, sostanze chimiche citotossiche o proteine citotossiche. Questa proprietà risulta valida per l'imaging dei tumori e nella terapia del cancro. Il rilascio controllato e mirato dei farmaci citotossici alle cellule tumorali limita il rischio di effetti collaterali nelle cellule normali (15).
Riepilogo
I numerosi microcompartimenti batterici si differenziano per natura e funzione, sono costituiti da gusci proteici e possono contenere proteine ed enzimi. Il rivestimento proteico di questi biocomposti agisce come una membrana agendo a livello della permeabilità del guscio BMC.
Attualmente esistono numerosi studi atti a comprendere ancor meglio la natura di queste proteine cellulari per poterle manipolarr utilizzando particolari forme di mutagenesi e per poterne modificare la porosità e la distribuzione della carica elettrica.
I biologi sintetici possono infatti così utilizzare questi microcompartimenti per creare gusci vuoti e trasportare i farmaci sul sito target.
Carboxysomes in Motion
Reference
Kerfeld et. al., 2018 Bacterial microcompartments. (79)
Turmo et. al., 2017 Carboxysomes: metabolic modules for CO2 fixation. 234(18).
Tanaka et. al., 2008 Atomic-level models of the bacterial carboxysome shell. 319(5866),1083-1086.
Chen et. al., 2012 Designing biological compartmentalization. 22(12),662-670.

Immagine CC0 Creative Commons, si ringrazia @mrazura per il logo ITASTEM.
CLICK HERE AND VOTE FOR DAVINCI.WITNESS
Please note that for organizational reasons it is necessary to use the "steemstem" and "davinci-times" tags to be voted.
@aditili
You got a 2.70% upvote from @getup
Want to promote your posts too? Send at least 0.010 STEEM DOLLAR or STEEM (max 0.055) to @getup with the post link as the memo and receive a upvote! More profits? Delegate some SteemPower to @getup - Daily Reward (STEEM DOLLAR)
1 SP, 5 SP, 10 SP, 100 SP, 500 SP, custom amount
► ► For Resteem to over 2000 follower + Upvote from @getup ◄ ◄
send 0.056 SBD with the post link as the memo.